Еще в 20-х гг. А. Лотка (Lotka), а несколько позднее независимо от него В. Вольтерра предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтерра. В основе модели лежит ряд допущений:
1) популяция жертв в отсутствие хищника растет экспоненциально,
2) пресс хищников тормозит этот рост,
3) смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций);
4) рождаемость хищника зависит от интенсивности потребления жертв.
Мгновенная скорость изменения численности популяции жертв может быть выражена уравнением
dN ж /dt = r 1 N ж - p 1 N ж N х,
где r 1 - удельная мгновенная скорость популяционного роста жертвы, p 1 - константа, связывающая смертность жертв с плотностью хищника, a N ж и N х - плотности соответственно жертвы и хищника.
Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости и постоянной смертности:
dN х /dt = p 2 N ж N х – d 2 N х,
где p 2 - константа, связывающая рождаемость в популяции хищника с плотностью жертв, a d 2 - удельная смертность хищника.
Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т.е. рост числа жертв лимитируется прессом хищников, а рост числа хищников - недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода популяции жертв из-под контроля хищника, хотя на самом деле такое бывает достаточно часто.
Несмотря на всю условность модели Лотки-Вольтерра, она заслуживает внимания уже хотя бы потому, что показывает, как даже такая идеализированная система взаимодействия двух популяций может порождать достаточно сложную динамику их численности. Решение системы этих уравнений позволяет сформулировать условия поддержания постоянной (равновесной) численности каждого из видoв. Популяция жертв сохраняет постоянную численность, если плотность хищника равна r 1 /p 1, а для того чтобы постоянство сохраняла популяция хищника, плотность жертв должна быть равна d 2 /p 2 . Если на графике отложить по оси абсцисс плотность жертв N ж , а по оси ординат - плотность хищника N х , то изоклины, показывающие условие постоянства хищника и жертвы, будут представлять собой две прямые, перпендикулярные друг другу и координатным осям (рис. 6,а). При этом предполагается, что ниже определенной (равной d 2 /p 2) плотности жертв плотность хищника всегда будет уменьшаться, а выше - всегда увеличиваться. Соответственно и плотность жертвы возрастает, если плотность хищника ниже значения, равного r 1 /p 1 , и уменьшается, если, она выше этого значения. Точка пересечения изоклин соответствует условию постоянства численности хищника и жертвы, а другие точки на плоскости этого графика совершают движение по замкнутым траекториям, отражая, таким образом, регулярные колебания численности хищника и жертвы (рис. 6,б). Размах колебаний определяется начальным соотношением плотностей хищника и жертвы. Чем ближе оно к точке пересечения изоклин, тем меньше окружность, описываемая векторами, и соответственно меньше амплитуда колебаний.
Рис. 6. Графическое выражение модели Лотки-Вольтера для системы хищник-жертва.
Одна из первых попыток получения колебаний численности хищника и жертвы в лабораторных экспериментах принадлежала Г.Ф. Гаузе (Gause). Объектами этих экспериментов были инфузория парамеция (Paramecium caudatum ) и хищная инфузория дидиниум (Didinium nasutum ). Пищей для парамеции служила регулярно вносимая в среду взвесь бактерий, а дидиниум питался только парамециями. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности приводил к полному истреблению жертв, после чего вымирала и популяция самого хищника. Усложняя опыты, Гаузе устраивал убежище для жертвы, внося в пробирки с инфузориями немного стеклянной ваты. Среди нитей ваты могли свободно перемещаться парамеции, но не могли дидиниумы. В таком варианте опыта дидиниум съедал всех парамеций, плавающих в свободной от ваты части пробирки, и вымирал, а популяция парамеции затем восстанавливалась за счет размножения особей, уцелевших в убежище. Некоторого подобия колебаний численности хищника и жертвы Гаузе удалось добиться только в том случае, когда он время от времени вносил в культуру и жертву и хищника, имитируя таким образом иммиграцию.
Через 40 лет после работы Гаузе его опыты были повторены Л. Лакинбиялом (Luckinbill), использовавшим в качестве жертвы инфузорию Paramecium aurelia , а в качестве хищника того же Didinium nasutum . Лакинбиллу удалось получить несколько циклов колебаний численности этих популяций, но только в том случае, когда плотность парамеций была лимитирована нехваткой пищи (бактерий), а в культуральную жидкость добавляли метилцеллюлозу - вещество, снижающее скорость движения как хищника, так и жертвы и потому уменьшающее частоту их возможных встреч. Оказалось также, что добиться колебаний хищника и жертвы легче, если увеличить объем экспериментального сосуда, хотя условие пищевого лимитирования жертвы и в этом случае обязательно. Если же к системе сосуществующих в колебательном режиме хищника и жертвы добавляли избыточную пищу, то ответом был, быстрый рост численности жертвы, за которым следовало возрастание численности хищника, приводящее в свою очередь к полному истреблению популяции жертвы.
Модели Лотки и Вольтерры послужили толчком для разработки ряда других более реалистичных моделей системы хищник-жертва. В частности, довольно простая графическая модель, анализирующая соотношение разных изоклин жертвы хищника, была предложена М. Розенцвейгом и Р. Мак-Артуров (Rosenzweig, MacArthur). Согласно этим авторам, стационарная (= постоянная) численность жертвы в координатных осях плотности хищника и жертвы может быть представлена в виде выпуклой изоклины (рис. 7,а). Одна точка пересечения изоклины сосью плотности жертвы соответствует минимальной допустимой плотности жертвы (нижеее популяция подвержена очень большому риску вымирания хотя бы из-за малой частоты встреч самцов и самок), а другая - максимальной, определяемой количеством имеющейся пищи или поведенческими особенностями самой жертвы. Подчеркнем, что речь идет пока о минимальной и максимальной плотностях в отсутствие хищника. При появлении хищника и увеличении его численности минимальная допустимая плотность жертвы, очевидно, должна быть выше, а максимальная - ниже. Каждому значению плотности жертвы должна соответствовать некоторая плотность хищника, при которой достигается постоянство популяции жертвы. Геометрическое место таких точек и есть изоклина жертвы в координатах плотности хищника и жертвы. Векторы, показывающие направление изменения плотности жертвы (ориентированные горизонтально), имеют разную направленность по разные стороны от изоклины (рис. 7,а).

Рис. 7. Изоклины стационарных популяций жертвы (а) и хищника (б).
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 7,б. определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.

Рис. 8. Возникновение колебательных режимов в системе хищник-жертва в зависимости от расположения изоклин хищника и жертвы.
При совмещении изоклин жертвы и хищника на одном, графике возможны три различных варианта (рис. 8). Если изоклина хищника пересекает изоклину жертвы в том месте, где она уже снижается (при высокой плотности жертв), векторы, показывающие изменение численности хищника и жертвы, образуют траекторию, закручивающуюся во внутрь, что соответствует затухающим колебаниям численности жертвы и хищника (рис. 8,а). В том случае, когда изоклина хищника пересекает изоклину жертвы в ее восходящей части (т.е. в области низких значений плотности жертв), векторы образуют раскручивающуюся траекторию, а колебания численности хищника и жертвы происходят соответственно с возрастающей амплитудой (рис. 8,б). Если же изоклина хищника пересекает изоклину жертвы в области ее вершины, то векторы образуют замкнутый круг, а колебания численности жертвы и хищника характеризуются стабильной амплитудой и периодом (рис. 8,в).
Иными словами, затухающие колебания соответствуют ситуации, при которой хищник ощутимо воздействует на популяцию жертв, достигнувшую только очень высокой плотности (близкой к предельной), а колебания возрастающей амплитуды возникают, тогда, когда хищник способен быстро увеличивать свою численность даже при невысокой плотности жертв и таким образом быстро ее уничтожить. В других вариантах своей модели Poзенцвейг и Мак-Артур показали, что стабилизировать колебания хищник-жертва можно, введя «убежище», т.е. предположив, что в области низкой плотности жертв существует область, где численность жертвы растет независимо от количества имеющихся хищников.
Стремление сделать модели более реалистичными путем их усложнения проявилось в работах не только теоретиков, но и экспериментаторов. В частности, интересные результаты были получены Хаффейкером (Huffaker), показавшим возможность сосуществования хищника и жертвы в колебательном режиме на примере мелкого растительноядного клеща Eotetranychus sexmaculatus и нападающего на него хищного клеща Typhlodromus occidentalis . В качестве пищи для растительноядного клеща использовали апельсины, помещенные на подносы с лунками (вроде тех, что используются для хранения и перевозки яиц). В первоначальном варианте на одном подносе было 40 лунок, причем в некоторых из них находились апельсины (частично очищенные от кожуры), а в других - резиновые мячики. Оба вида клещей размножаются партеногенетически очень быстро, и поэтому характер их популяционной динамики можно выявить за сравнительно короткий срок. Поместив на поднос 20 самок растительноядного клеща Хаффейкер наблюдал быстрый рост его популяции, которая стабилизировалась на уровне 5-8 тыс. особей (в расчете на один апельсин). Если к растущей популяции жертвы добавляли несколько особей хищника, то популяция последнего быстро увеличивала свою численность и вымирала, когда все жертвы оказывались съеденными.
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом 6 апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 месяцев, продемонстрировав три полных цикла колебаний численности. Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum ), завезенная в Австралию, значительно снизила численность кактуса опунции, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию.
Говоря о колебаниях хищник-жертва, нельзя не упомянуть и о циклических изменениях численности зайца и рыси в Канаде, прослеженных по материалам статистики заготовок пушнины компанией Гудзон-Бэй с конца XVIII вплоть до начала XX в. Этот пример нередко рассматривался как классическая иллюстрация колебаний хищник-жертва, хотя на самом деле мы видим только следование роста численности популяции хищника (рыси) за ростом численности жертвы (зайца). Что же касается снижения численности зайцев после каждого подъема, то оно не могло объясняться только возросшим прессом хищников, а было связано с другими факторами, по-видимому, прежде всего нехваткой корма в зимний период. К такому выводу пришел, в частности, М. Джилпин (Gilpin), пытавшийся проверить, могут ли быть описаны эти данные классической моделью Лотки-Вольтерры. Результаты проверки показали, что удовлетворительного соответствия модели нет, но как ни странно, оно становилось лучше, если хищника и жертву меняли местами, т.е. трактовали рысь как «жертву», а зайца - как «хищника». Подобная ситуация нашла свое отражение и в шутливом названии статьи («Едят ли зайцы рысей?»), по сути своей очень серьезной и опубликованной в серьезном научном журнале.
Еще в 20-х гг. А. Лотка, а несколько позднее независимо от него В.Вольтера предложили математические модели, описывающие сопряженные колебания численности хищника и жертвы.
Модель состоит из двух компонентов:
С – численность хищников; N – численность жертв;
Предположим, что в отсутствии хищников популяция жертвы будет расти экспоненциально: dN/dt = rN. Но жертвы уничтожаются хищниками со скоростью, которая определяется частотой встреч хищника с жертвой, а частота встреч возрастает по мере увеличения численности хищника (С) и жертвы (N). Точное число встреченных и успешно съеденных жертв будет зависеть от эффективности, с которой хищник находит и ловит жертву, т.е. от а’ – «эффективности поиска» или «частоты нападений». Т.о., частота «успешных» встреч хищника с жертвой и, следовательно, скорость поедания жертв будет равна а’СN и в целом: dN/dt = rN – a’CN (1*).
В отсутствии пищи отдельные особи хищника теряют в весе, голодают и гибнут. Предположим, что в рассматриваемой модели численность популяции хищника в отсутствии пищи вследствие голодания будет уменьшаться экспоненциально: dC/dt = - qC, где q – смертность. Гибель компенсируется рождением новых особей со скоростью, которая, как полагают в этой модели, зависит от двух обстоятельств:
1) скорости потребления пищи, a’CN;
2)эффективности (f), с которой эта пища переходит в потомство хищника.
Т.о., рождаемость хищника равна fa’CN и в целом: dC/dt = fa’CN – qC (2*). Уравнения 1* и 2* составляют модель Лотки-Вольтера. Свойства этой модели можно исследовать, построить изоклины линии, соответствующие постоянной численности популяции, с помощью таких изоклин определяют поведение взаимодействующих популяций хищник-жертва.
В случае с популяцией жертвы: dN/dt = 0, rN = a’CN, или C = r/a’. Т.к. r и a’ = const, изоклиной для жертвы будет линия, для которой величина С является постоянной:
При низкой плотности хищника (С), численность жертвы (N) возрастает, наоборот – снижается.
Точно также для хищников (уравнение 2*) при dC/dt = 0, fa’CN = qC, или N = q/fa’, т.е. изоклиной для хищника будет линия, вдоль которой N постоянна: При высокой плотности жертвы, численность популяции хищника растет, а при низкой – снижается.
Их численность претерпевает неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищников на популяцию жертвы и тем самым к уменьшению ее численности. Это снижение в свою очередь ведет к ограничению хищников в пище и падению их численности, которая вызывает ослабление пресса хищников и увеличение численности жертвы, что снова приводит к росту популяции хищников и т.д.
Популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новые циклы неограниченных колебаний. На самом деле среда постоянно меняется, и численность популяций будет постоянно смещаться на новый уровень. Чтобы циклы колебаний, которые совершает популяция, были регулярными, они должны быть стабильными: если внешнее воздействие изменяет уровень численности популяций, то они должны стремиться к первоначальному циклу. Такие циклы называются устойчивыми, предельными циклами.
Модель Лотки-Вольтера позволяет показать основную тенденцию в отношениях хищник-жертва, которая выражается в возникновении колебаний численности в популяции жертвы, сопровождающихся колебаниями численности в популяции хищника. Основным механизмом таких колебаний является запаздывание во времени, свойственное последовательности состояния от высокой численности жертв к высокой численности хищников, затем к низкой численности жертв и низкой численности хищников, к высокой численности жертв и т.д.
5) ПОПУЛЯЦИОННЫЕ СТРАТЕГИИ ХИЩНИКА И ЖЕРТВЫ
Взаимоотношения «хищник - жертва» представляют звенья процесса передачи вещества и энергии от фитофагов к зоофагам или от хищников низшего порядка к хищникам высшего порядка. По характеру этих отношений различают три варианта хищников :
а) собиратели . Хищник собирает мелких достаточно многочисленных подвижных жертв. Такой вариант хищничества характерен для многих видов птиц (ржанок, зябликов, коньков и др.), которые затрачивают энергию только на поиск жертв;
б) истинные хищники . Хищник преследует и убивает жертву;
в) пастбищники . Эти хищники используют жертву многократно, например, оводы или слепни.
Стратегия добывания пищи у хищников направлена на обеспечение энергетической эффективности питания: затраты энергии на добывание пищи должны быть меньше энергии, получаемой при ее усвоении.
Истинные хищники делятся на
«жнецов», которые питаются обильными ресурсами (н, планктонные рыбы и даже усатый кит), и «охотников», которые добывают менее обильный корм. В свою очередь
«охотники» делятся на «засадников», подстерегающих добычу (например, щука, ястреб, кошка, жук- богомол), «искателей» (насекомоядные птицы) и «преследователей». Для последней группы поиск пищи не требует больших затрат энергии, но ее нужно много, чтобы овладеть жертвой (львы в саваннах). Впрочем, некоторые хищники могут сочетать элементы стратегии разных вариантов охоты.
Как и при отношениях «фитофаг - растение», ситуация, при которой все жертвы будут съедены хищниками, что в конечном итоге приведет и к их гибели, в природе не наблюдается. Экологическое равновесие между хищниками и жертвами поддерживается специальными механизмами , снижающими риск полного истребления жертв. Так, жертвы могут :
Убегать от хищника. В этом случае в результате адаптаций повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей;
Приобретать защитную окраску («притворяться» листьями или сучками) или, напротив, яркий цвет, Н.: красный, предупреждающий хищника о горьком вкусе. Общеизвестно изменение окраски зайца в разные времена года, что позволяет ему маскироваться летом в траве, а зимой на фоне белого снега. Адаптивное изменение окраски может происходить в разных стадиях онтогенеза: детеныши тюленей белые (цвет снега), а взрослые особи черные (цвет скалистого побережья);
Распространяться группами, что делает их поиск и промысел для хищника более энергоемким;
Прятаться в укрытия;
Переходить к мерам активной обороны (травоядные, имеющие рога, колючие рыбы), иногда совместной (овцебыки могут занимать «круговую оборону» от волков и т.д.).
В свою очередь хищники развивают не только способность к быстрому преследованию жертв, но и обоняние, позволяющее по запаху определить место нахождения жертвы. Многие виды хищников разрывают норы своих жертв (лисы, волки).
В то же время сами они делают все возможное для того, чтобы не обнаружить своего присутствия. Этим объясняется чистоплотность мелких кошачьих, которые для устранения запаха много времени тратят на туалет и закапывают экскременты. Хищники одевают «маскировочные халаты» (полосатость щук и окуней, делающих их менее заметными в зарослях макрофитов, полосатость тигров и т.д.).
Полной защиты от хищников всех особей в популяциях животных-жертв также не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге - к катастрофе популяций жертв. В то же время при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные) и ввиду резкого увеличения их численности подрывается кормовая база.
По этой причине эффект зависимости численности популяций жертв и хищников - пульсация численности популяции жертвы, за которой с некоторым запаздыванием пульсирует численность популяции хищника («эффект Лотки - Вольтерры») - наблюдается редко.
Между биомассами хищников и жертв устанавливается достаточно устойчивое соотношение. Так, Р. Риклефс приводит данные о том, что соотношение биомасс хищника и жертвы колеблется в пределах 1:150 - 1:300. В разных экосистемах умеренной зоны США на одного волка приходится 300 мелких белохвостых оленей (вес 60 кг), 100 крупных оленей вапити (вес 300 кг) или 30 лосей (вес 350). Такая же закономерность выявлена в саваннах.
При интенсивной эксплуатации популяций фитофагов человек нередко исключает из экосистем хищников (в Великобритании, к примеру, есть косули и олени, но нет волков; в искусственных водоемах, где разводят карпа и другую прудовую рыбу, нет щук). В этом случае роль хищника выполняет сам человек, изымая часть особей популяции фитофага.
Особый вариант хищничества наблюдается у растений и грибов. В царстве растений встречается порядка 500 видов, способных ловить насекомых и частично их переваривать с помощью протеолитических ферментов. Хищные грибы образуют ловчие аппараты в виде маленьких овальных или шаровидных головок, расположенных на коротких веточках мицелия. Однако самый распространенный вид ловушки - клейкие трехмерные сети, состоящие из большого числа колец, образующихся в результате ветвления гиф. Хищные грибы могут ловить достаточно крупных животных, например, круглых червей. После того, как червь запутается в гифах, они прорастают внутрь тела животного и быстро его заполняют.
1.Постоянный и благоприятный уровень температуры и влажности.
2.Обилие пищи.
3.Защита от неблагоприятных факторов.
4.Агрессивный химический состав среды обитания (пищеварительные соки).
1.Наличие двух сред обитания: среда первого порядка - организм хозяина, среда второго порядка - внешняя среда.
Федеральное агентство по образованию
Государственное образовательное учреждение
высшего профессионального образования
«Ижевский государственный технический университет»
Факультет «Прикладная математика»
Кафедра «Математическое моделирование процессов и технологий»
по дисциплине «Дифференциальные уравнения»
Тема: «Качественное исследование модели хищник-жертва»
Ижевск 2010
ВВЕДЕНИЕ
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.2 Обобщенные модели Вольтера типа «хищник-жертва».
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
В настоящее время задачи экологии имеют первостепенное значение. Важным этапом решения этих задач является разработка математических моделей экологических систем.
Одной из основных задач экологии па современном этапе является изучение структуры и функционирования природных систем, поиск общих закономерностей. Большое влияние на экологию оказала математика, способствующая становлению математической экологии, особенно такие её разделы, как теория дифференциальных уравнений, теория устойчивости и теория оптимального управления.
Одной из первых работ в области математической экологии была работа А.Д. Лотки (1880 - 1949), который первый описал взаимодействие различных популяций, связанных отношениями хищник - жертва. Большой вклад в исследование модели хищник -жертва внесли В. Вольтерра (1860 - 1940), В.А. Костицин (1883-1963) В настоящее время уравнения описывающие взаимодействие популяций, называются уравнениями Лотки - Вольтерра.
Уравнения Лотки - Вольтерра описывают динамику средних величин - численности популяции. В настоящее время на их основе построены более общие модели взаимодействия популяций, описываемые интегро-дифференциальными уравнениями, исследуются управляемые модели хищник - жертва.
Одной из важных проблем математической экологии является проблема устойчивости экосистем, управления этими системами. Управление может осуществляться с целью перевода системы из одного устойчивого состояния в другое, с целью её использования или восстановления.
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Попытки математического моделирования динамики как отдельных биологических популяций, так и сообществ, включающих взаимодействующие популяции различных видов, предпринимались давно. Одна из первых моделей роста изолированной популяции (2.1) была предложена еще в 1798 г. Томасом Мальтусом:
, (1.1)Данная модель задается следующими параметрами:
N - численность популяции;
- разность между коэффициентами рождаемости и смертности.Интегрируя это уравнение получаем:
, (1.2)где N(0) – численность популяции в момент t = 0. Очевидно, что модель Мальтуса при
> 0 дает бесконечный рост численности, что никогда не наблюдается в природных популяциях, где ресурсы, обеспечивающие этот рост, всегда ограничены. Изменения численности популяций растительного и животного мира нельзя описывать простым законом Мальтуса, на динамику роста влияют многие взаимосвязанные причины – в частности, размножение каждого вида саморегулируется и видоизменяется так, чтобы этот вид сохранялся в процессе эволюции.Математическим описанием этих закономерностей занимается математическая экология – наука об отношениях растительных и животных организмов и образуемых ими сообществ между собой и с окружающей средой.
Наиболее серьезное исследование моделей биологических сообществ, включающих в себя несколько популяций различных видов, было проведено итальянским математиком Вито Вольтерра:
, - численность популяции; - коэффициенты естественного прироста (или смертности) популяции; - коэффициенты межвидового взаимодействия. В зависимости от выбора коэффициентов модель описывает либо борьбу видов за общий ресурс, либо взаимодействие типа хищник - жертва, когда один вид является пищей для другого. Если в работах других авторов основное внимание уделялось построению различных моделей, то В. Вольтерра провел глубокое исследование построенных моделей биологических сообществ. Именно с книги В. Вольтерра, по мнению многих ученых, началась современная математическая экология.2. КАЧЕСТВЕННОЕ ИССЛЕДОВАНИЕ ЭЛЕМЕНТАРНОЙ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.1 Модель трофического взаимодействия по типу «хищник-жертва»
Рассмотрим модель трофического взаимодействия по типу «хищник-жертва», построенную В. Вольтерром. Пусть имеется система, состоящая из двух видов, из которых один поедает другой.
Рассмотрим случай, когда один из видов является хищником, а другой - жертвой, и будем считать, что хищник питается только жертвой. Примем следующую простую гипотезу:
- коэффициент прироста жертвы; - коэффициент прироста хищника; - численность популяции жертвы; - численность популяции хищника; - коэффициент естественного прироста жертвы; - скорость потребления жертвы хищником; - коэффициент смертности хищника в отсутствие жертвы; - коэффициент «переработки» хищником биомассы жертвы в собственную биомассу.Тогда динамика численности популяций в системе хищник - жертва будет описываться системой дифференциальных уравнений (2.1):
(2.1)где все коэффициенты положительные и постоянные.
Модель имеет равновесное решение (2.2):
(2.2)По модели (2.1) доля хищников в общей массе животных выражается формулой (2.3):
(2.3)Анализ устойчивости состояния равновесия по отношению к малым возмущениям показал, что особая точка (2.2) является «нейтрально» устойчивой (типа «центр»), т. е. любые отклонения от равновесия не затухают, но переводят систему в колебательный режим с амплитудой, зависящей от величины возмущения. Траектории системы на фазовой плоскости
имеют вид замкнутых кривых, расположенных на различных расстояниях от точки равновесия (рис. 1).Рис. 1 – Фазовый «портрет» классической вольтерровой системы «хищник-жертва»
Разделив первое уравнение системы (2.1) на второе, получим дифференциальное уравнение (2.4) для кривой на фазовой плоскости
. (2.4)Интегрируя данное уравнение получим:
(2.5) - постоянная интегрирования, гдеНесложно показать, что движение точки по фазовой плоскости будет происходить только в одну сторону. Для этого удобно сделать замену функций
и , перенеся начало координат на плоскости в стационарную точку (2.2) и введя затем полярные координаты: (2.6)В таком случае, подставив значения системы (2.6) в систему (2.1), будем иметь.
Модель ситуации типа «хищник-жертва»
Рассмотрим математическую модель динамики сосуществования двух биологических видов (популяций), взаимодействующих между собой по типу «хищник-жертва» (волки и кролики, щуки и караси и т.д.), называемую моделью Волътера-Лотки. Впервые она была получена А. Лоткой (1925 г.), А чуть позже и независимо от Лотки аналогичные и более сложные модели были разработаны итальянским математиком В. Вольтерра (1926 г.), работы которого фактически заложили основы так называемой математической экологии.
Пусть есть два биологических вида, которые совместно обитают в изолированной среде. При этом предполагается:
- 1. Жертва может найти достаточно пищи для пропитания;
- 2. При каждой встрече жертвы с хищником последний убивает жертву.
Будем для определённости называть их карасями и щуками. Пусть
состояние системы определяется величинами x(t) и y(t) - количеством карасей и щук в момент г. Чтобы получить математические уравнения, которые приближенно описывают динамику (изменение во времени) популяции, поступим следующим образом.
Как и в предыдущей модели роста популяции (см. п. 1.1) для жертв имеем уравнение
где а > 0 (рождаемость превышает смертность)
Коэффициент а прироста жертв зависит от количества хищников (уменьшается с их увеличением). В простейшем случае а- а - fjy (а>0,р>0). Тогда для численности популяции жертв имеем дифференциальное уравнение
Для популяции хищников имеем уравнение

где b >0 (смертность превышает рождаемость).
Коэффициент b убывания хищников уменьшается, если имеются жертвы, которыми можно питаться. В простейшем случае можно принять b - у -Sx {у > 0, S > 0). Тогда для численности популяции хищников получим дифференциальное уравнение

Таким образом, уравнения (1.5) и (1.6) представляют собой математическую модель рассматриваемой задачи взаимодействия популяций. В этой модели переменные х,у - состояние системы, а коэффициенты характеризуют ее структуру. Нелинейная система (1.5), (1.6) и есть модель Вольтера-Лотки.
Уравнения (1.5) и (1.6) следует дополнить начальными условиями - заданными значениями начальных популяций.
Проведем теперь анализ построенной математической модели.
Посгроим фазовый портрет системы (1.5), (1.6) (по смыслу задачи х > 0, v >0). Разделив уравнение (1.5) на уравнение (1.6), получим уравнение с разделяющимися переменными
Игшлрируя это уравнение, будем иметь
Соотношение (1.7) даёт уравнение фазовых траекторий в неявном виде. Система (1.5), (1.6) имеет стационарное состояние определяемое из

Из уравнений (1.8) получим (т.к. л* Ф 0, у* Ф 0)
Равенства (1.9) определяют на фазовой плоскости положение равновесия (точку О) (Рис. 1.6).

Направление движения по фазовой траектории можно определить из таких соображений. Пусть карасей мало. г.е. х ~ 0, тогда из уравнения (1.6) у
Все фазовые траектории (за исключением точки 0) замкнутые кривые, охватывающие положение равновесия. Состоянию равновесия соответствует неизменное количество х« и у« карасей и щук. Караси размножаются, щуки их едят, вымирают, но число тех и дрч их не меняется. "Замкнутым фазовым траекториям соответствует периодическое изменение численности карасей и щук. Причём то, по какой траектории движется фазовая точка, зависит от начальных условий. Рассмотрим, как меняется состояние вдоль фазовой траектории. Пусть точка находится в положении А (рис. 1.6). Здесь карасей мало, щук много; щукам есть нечего, и они постепенно вымирают и почти
совсем исчезают. Но и количество карасей тоже уменьшается почти до нуля и
только потом, когда щук стало меньше, чем у , начинается прирост количества карасей; скорость их прироста увеличивается и их число увеличивается - так происходит примерно до точки В. Но увеличение числа карасей приводит к торможению процесса вымирания шук и их число начинает расти (пищи стало больше) - участок ВС. Далее щук много, они едят карасей и почти всех съедают (участок CD). После этого щуки снова начинают вымирать и процесс повторяется с периодом примерно в 5-7 лет. На рис. 1.7 качественно построены кривые изменения численности карасей и щук в зависимости от времени. Максимумы кривых чередуются, причём максимумы численности щук отстают от максимумов карасей.

Такое поведение характерно для различных систем типа хищник - жертва. Проведем теперь интерпретацию полученных результатов.
Несмотря на то, что рассмотренная модель является простейшей и в действительности всё происходит гораздо сложнее, она позво.чила объяснить кое-что из загадочного, чго есть в природе. Понятны рассказы рыболовов о периодах, когда «щуки сами прыгают в руки», получила объяснение периодичность протекания хронических болезней и т.д.
Отметим еще один интересный вывод, который можно сделать из Рис. 1.6. Если в точке Р происходит быстрый отлов щук (в другой терминологии - отстрел волков), то система «перепрыгивает » в точку Q, и дальнейшее движение происходит по замкнутой траектории меньшего размера, что интуитивно ожидаемо. Если же уменьшить число щук в точке R, то система перейдет в точку S, и дальнейшее движение будет происходить по траектории большего размера. Амплитуды колебаний увеличатся. Это противоречит интуиции, но как раз объясняет такое явление: в результате отстрела волков их численность увеличивается со временем. Таким образом, важным в этом случае является выбор момента отстрела.
Предположим, что две популяции насекомых (например, тля и божья коровка, которая есть тлю) находились в естественном равновесии х-х*,у = у* (точка О на Рис. 1.6). Рассмотрим влияние разового применения инсектицида, который убивает х> 0 из жертв и у > 0 из хищников, не уничтожая их полностью. Уменьшение численности обеих популяций приводит к тому, что изображающая точка из положения О «перескочит» ближе к началу координат, где х > 0, у 0 (Рис. 1.6) Отсюда следует, что в результате действия инсектицида, призванного уничтожать жертв (тлю), число жертв (тли) увеличивается, а число хищников (божьих коровок) уменьшается. Получается, что численность хищников может стать настолько малой, что им будет фозить полное исчезновение но другим причинам (засуха, болезни и т.д.). Таким образом, применение инсектицидов (если только они не уничтожают вредных насекомых практически полностью) в конечном счёте приводит к увеличению популяции тех насекомых, численность которых находилась под контролем других насекомых-хищников. Такие случаи описаны в книгах по биологии.
В общем случае коэффициент прироста количества жертв а зависит и от Л" и от у: а = а(х, у) (из-за наличия хищников и из-за ограничений на пищу).
При малом изменении модели (1.5), (1.6) к правым частям уравнений добавляются малые члены (учитывающие, например, конкуренцию карасей за пищу и щук за карасей)
здесь 0 f.i « 1.
В таком случае вывод о периодичности процесса (возвращении системы к исходному состоянию), справедливый для модели (1.5), (1.6), теряет силу. В зависимости от вида малых поправок/ и g возможны ситуации, показанные на Рис. 1.8.

В случае (1) равновесное состояние О устойчиво. При любых других начальных условиях через достаточно большое время устанавливается именно оно.
В случае (2) система «идёт в разнос». Стационарное состояние неустойчиво. Такая система в конце концов попадает в такую область значений х и у, что модель перестаёт быть применимой.
В случае (3) в системе с неустойчивым стационарным состоянием О устанавливается с течением времени периодический режим. В отличие от исходной модели (1.5), (1.6) в этой модели установившийся периодический режим не зависит от начальных условий. Первоначально малое отклонение от стационарного состояния О приводит не к малым колебаниям около О , как в модели Вольтерра-Лотки, а к колебаним вполне определённой (и не зависящей от малости отклонения) амплитуды.
В.И. Арнольд называет модель Вольтерра-Лотки жесткой, т.к. её малое изменение может привести к выводам, отличным от приведенных выше. Для суждения о том, какая из ситуаций, указанных на Рис. 1.8, реализуется в данной системе, совершенно необходима дополнительная информация о системе (о виде малых поправок/ и g ).
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
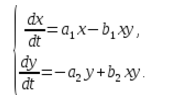
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).